lncRNA是一类长度大于200个核苷酸的RNA分子。根据lncRNA相对于附近编码基因的位置将其分为antisense、intronic、intergenic、bidirectional和启动子相关等5类lncRNA,这些lncRNA对编码基因的调控方式大有不同。antisense lncRNA是一类与正向基因具有部分外显子重叠的反义RNA,这种天然反义lncRNA的存在及位置的特殊性揭示其与正义链的作用往往比一般靶基因更密切。那么,antisense lncRNA作为数量较多的一类lncRNA,它是如何发挥作用的呢?下面从表观遗传学、转录及转录后水平进行简单介绍。
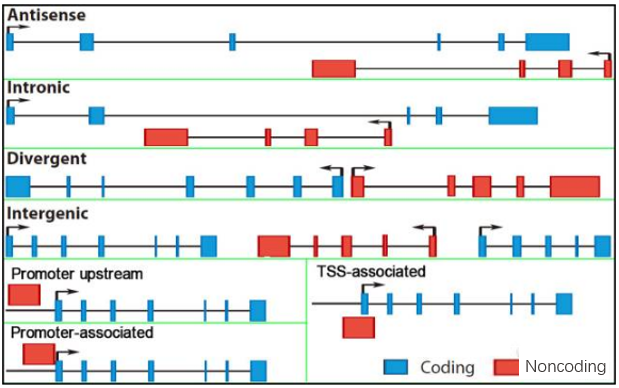
图1 lncRNA相对于编码基因位置
表观遗传调控
在不改变基因核苷酸序列的情况下,antisense lncRNA能够结合DNA进行甲基化酶及染色质组蛋白修饰酶的招募,通过改变染色质状态,调控正义基因的表达,并且产生可遗传变异。像这种修饰可以小到作用于一个基因,大到影响整个染色体,比如Xist和反义Tsix二者介导的X染色体失活过程。
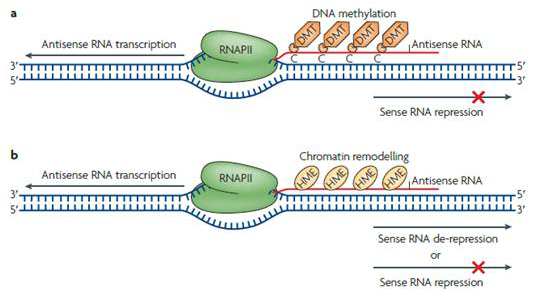
图2 antisense lncRNA产生的表观遗传修饰
由于哺乳动物雌性含有两条X染色体,而雄性只有一条X染色体,为了确保雄性(XY)和雌性(XX)的X基因剂量相等,雌性胚胎会发生单个X染色体永久失活,即“剂量补偿”效应。非编码RNA Xist是X染色体失活的主要参与基因,Xist仅在失活的X染色体中表达,其通过包裹合成它的X染色体,并结合Eed/PRC2复合物形成H3K27me3组蛋白修饰,进而抑制基因转录,致使X染色体失活。而在Xist基因3'端存在antisense lncRNA Tsix,Tsix的转录可以阻止Eed/PRC2募集到Xist启动子区并且抑制H3K4甲基化,导致染色质结构发生改变,最终沉默Xist。
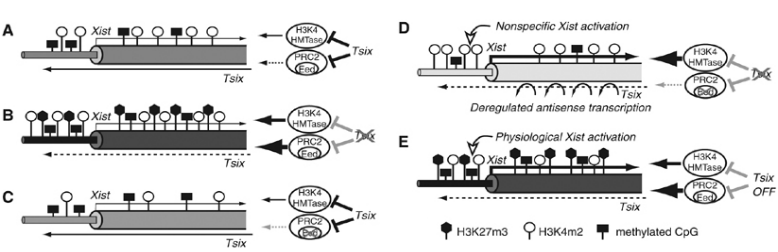
图3 Tsix对Xist染色质的修饰状态1
转录水平
由于antisense lncRNA和正义链基因方向相反,单单从空间位置来说,有模型表明当正/反义链同时转录形成RNA时,RNA聚合酶会相遇产生一定的空间阻碍,从而使RNA的转录受到抑制,像这种转录调控是由于基因本身方向相反导致,并不是由antisense lncRNA本身的调控功能引起对正义链基因转录的影响。
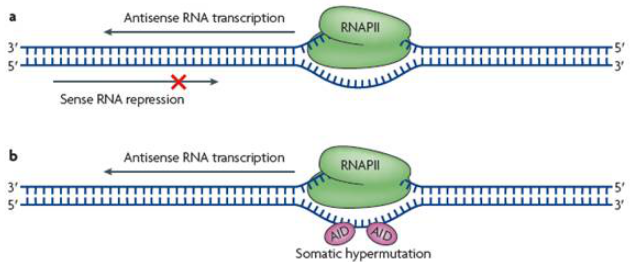
图4 antisense lncRNA转录模式图2
在细胞核内,antisense lncRNA能够与正义链RNA通过碱基配对形成二聚体,二聚体的形成会导致部分外显子的剪接位点被掩盖掉,从而影响前体pre-mRNA的剪切过程,产生不同种类的mRNA成熟体。研究表明,antisense lncRNA除了通过二聚体形式调控pre-mRNA的成熟剪切,它也可以对核内mRNA的部分核苷酸进行替换编辑,如果蝇的反义Sas-10通过结合Rnp4f,将Rnp4f部分腺嘌呤替换为次黄嘌呤,从而降低Rnp4f mRNA的表达水平。
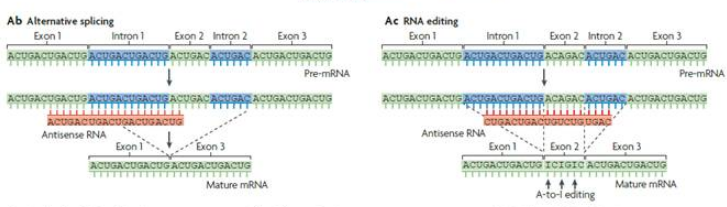
图5 核内正/反义链RNA 碱基配对形成二聚体形式2
转录后水平
Antisense lncRNA不只在细胞核内影响正义基因的转录,正/反义链RNA二聚体同样能够在转录后水平调控基因的表达。antisense lncRNA可以结合正义基因mRNA 3’ UTR区,封闭大量miRNA的靶位点,阻碍了miRNA对靶基因的抑制作用,从而维持mRNA的稳定性。如反义lncRNA-PXN-AS1-L能够结合PXN mRNA 3’ UTR,提高PXN mRNA的稳定性,最终促进癌基因RXN的表达和肿瘤细胞的生长3。
当然,antisense lncRNA同样能够吸附miRNA,进而减弱miRNA对靶基因的下调。另外,antisense lncRNA可与编码蛋白基因的转录本形成互补双链,在Dicer酶的作用下产生类似内源性的siRNA,从而沉默编码基因的表达。
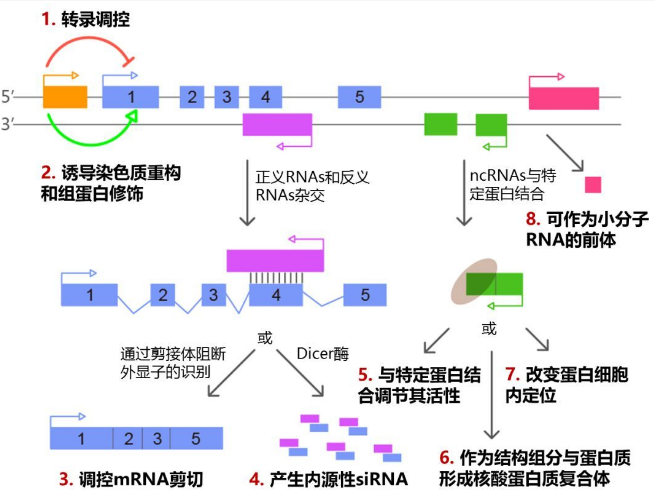
图6 lncRNA作用机制
目前很多关于antisense lncRNA的调控机制不是很清楚,但很多研究表明其在机体的生长、发育和疾病发生过程中起着重要作用。所以,只要我们锲而不舍深入挖掘,总会有机制明了的时候。
吉凯基因具有高效的质粒、病毒等工具表达载体,可以实现非编码基因在细胞及动物水平上的过表达、敲低和敲除等目的,为非编码RNA调控机制的研究提供便利,同时为广大优秀的科研学者助力。

关注 | 病毒实验帮
公众号底部菜单栏【新功能】上线!
免费在线学习《国自然热点研究》、《数据库及软件操作教程》
一键下载《病毒使用手册》、《高分文献》